
生命、物理和数学定律的回顾
(http://www.prequark.org/Overview.htm )
Copyright © 1994 by Tienzen (Jeh-Tween) Gong
Ⅰ、导论
Ⅱ、细胞分化的初始信号 — 几何学
Ⅲ、生命的积极行动者 — 时间和空间
Ⅳ、过去、现在和将来之间的相互作用
Ⅴ、相互包容无处不在
Ⅵ、体形和大脑结构的规律
Ⅶ、对称性及其破缺
Ⅰ、导论
看看现在所有多细胞的生命形式,产生了许多问题:
第一,从一个单一的受精卵(结合子),如何产生了这么多形式各异的细胞?
第二、当分裂的细胞想聚合在一起,形成更大的单元时,他们都放弃了自己的独立性,而移交了大部分的功能(如:繁殖力)。他们为什么想合作呢?这种合作(分工)又是如何开展的?
第三、不同的物种有着不同的形体,其形状是由基因控制的吗?或者,基因是由体形所创造的(作为创建特定体形的工具)?
第四、卵细胞(或者是任何器官)是受驱动这么做的吗?还是,它自己任凭意愿选择了命运?如果是受到驱动,那么驱动力何在?如果是凭着自己的意愿,那么,它又是如何找到生命的发展程式的?
第五、每一个生命的发展形式都在时间和空间的舞台上演绎着自己的角色。时空仅仅是舞台吗?或者,可能他们才是生命戏剧里的真正主演?
体型和基因之间的相互关系涉及了拓扑学,而发展和时空的关系则与物理学有关。可以肯定的是,驱动力和自由意愿都是某一种智力性的东西。这个过程包括了哲学、形而上学、数学、物理和神学。但不幸的是,迄今为止,对生命科学的定义只局限于胚胎学、分子生物学和遗传学的范围。由于在所谓的生命科学与其他基础学科(数学、物理和神学)之间没有绝对的联系(一种统一体系),对生命的真正理解还远远不够。事实上,对以上这些问题,哪怕你对基因学和生物学所知不多,也可以理解、提出并作出回答。本文旨在不仅用普通常识来提出这些问题(不需要分子生物学和基因学的知识),还要提供一个各个学科(数学、物理学、神学和生物学)之间的统一的体系。
Ⅱ、细胞分化的初始信号 — 几何
早期生命的发展可以分为三个阶段 :
1。 单个受精卵分裂形成为一个实心球(桑葚胚),在卵裂阶段形状象桑葚团。
2。 这个桑葚胚长成一个 {中空的球(芽状物的希腊语称为blastula)}。
3。 第三个阶段, 就是原肠胚形成阶段(这个阶段对任何个体来说都是最重要的)–胞胚细胞的前半部分形成一个软管(DONUT)。里层,即中胚叶形成了大部分的内部器官。外层即外胚层,形成了皮肤和中枢和周边神经系统。
從 桑葚胚 (morula) 到 Blastula
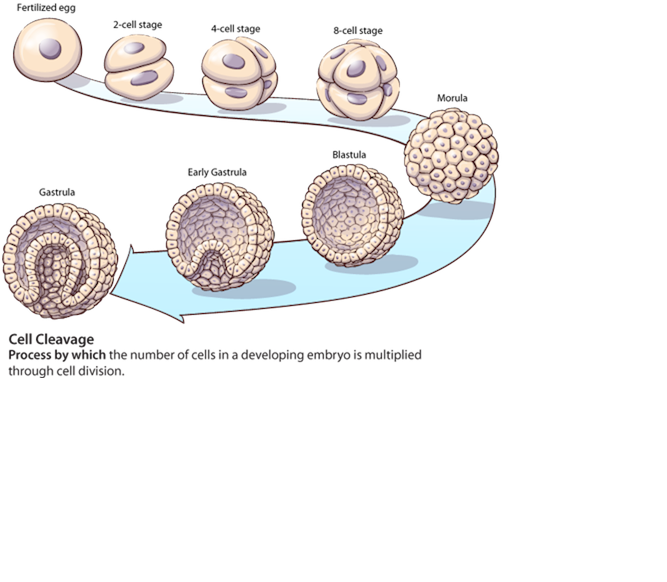
從 blastula 到 gastrulation 到 embryo (胎胞)
在卵裂阶段,所有细胞都是等同的。在果蝇实验中,将卵裂阶段的一个雄细胞引入雌性胚胎形成雌雄嵌体(雌雄各半的个体),而其中的各部分是随机分布的。也就是说,在这个阶段细胞的命运还是没有决定。
在胞胚阶段,细胞开始分化,而这个分化依赖于胞胚中各细胞的分布情况。其腹部就形成内部器官。如果在这个时候细胞移动到另外的地方,那么这种初始阶段的分化还是可逆的。胞胚的背部则在形成原肠胚时形成外层和神经系统。但是其确定的分化一定要等到他们与其他区域(中胚层)发生联系后才会开始,否则不会发生。
迄今为止,这种重要的生命过程,并不需要用分子生物学或基因学的特定术语来描绘。它只是几何的一种简单变形:从一个单细胞成为实心球(桑葚胚);从实心球到中空球(胞胚);从中空球到环形管(DONUT)。这是一种导致细胞分化的几何提法。几何关系赋予了细胞的命运。我的论文–How to reach Ultinate Reality in a finite number of steps in terms of sciences(如何用科学术语在有限步骤内达到终极实在)(发表在1993年莫斯科的19届全球哲学大会和1995年多伦多大学的关于终极实在和意义的人类思想研究国际学会的两年一度例会上)根据球-环形管的概念提供了一个拓扑学、物理学、数学和生命学间的统一体系。
Ⅲ、生命的积极行动者 — 时间和空间
在所有生命科学家(分子生物学家、基因学家)的眼里,在生命中演绎的主要角色是基因、酶、蛋白质,它们在空间和时间的舞台上,上演着生命的戏剧。没有人会亲眼看到过空间和时间的存在。
每个卵都有其特定的拓扑结构(几何结构),而且通常说来是非对称的。尽管许多不同的实验(受外力干扰)都可以改变胚胎的轴心(结束于头或尾),卵的形状确实是与轴心有一定关联。
果蝇的实验证明,其形体主要是由四个成形素的梯度所控制。成形素是控制分化模式的物质。头的基因(BICOID)必须要分布在生长脑袋的地方,而不是(至少不是高浓度)分布于其他地方。否则将会到处长满脑袋。该过程是通过下列方法实现的:
首先,是卵的极化,在一端产生头基因(BICOID),在另一端产生尾基因。
其次,如果可以允许头基因扩散到尾部,那么,在尾部将会产生一个脑袋。但是,头基因的半衰期还不到一个小时,而两端的巨大差异将阻止它到达尾部。
再者,尾部基因具有破坏头基因的能力,为了避免在腹部长出一个脑袋,即使是在卵的中间部分,头基因的浓度也会被破坏至极低的水平。
由此可知,果蝇(或其它多细胞生命体)的体形是由四要素决定的:
- 拓扑和化学极化结构;
- 每个器官的基因;
- 每个基因的半衰期;
- 基因之间的几何分化。
简言之,时间、空间和几何控制了生命的发展,而基因仅仅是蓝领工作者而已。
IV。 过去,现在和将来的相互作用
体型发展受控于当时的基因层次。在实时的层次中,有两类有否决权的基因。
*一是上游否决基因,它指定了下游基因的正确的位置。
*二是下游否决基因,它能驳回先前的行为。
受精卵中的性染色体,通过细胞的多代分裂传递给身体的所有细胞。因此,每个细胞,不管是在肝脏,肌肉或大脑中都有决定自身性别的信息,和整个生物体的性信息相同。然而,性决定基因TDF(Testis-Determining Factor 睾丸决定因子)只在一个组织中作用,即性腺中。在受精后的第五和第七周之间,任一性别的人类胚胎,都发展多用途的性腺,以后可以变成睾丸或卵巢.如果TDF(男性性基因)受到任何方式的阻碍或转化,性腺一直等待到第十三周,然后开始发展为卵巢,他就变成她了。
如果他有睾丸,在第八周如果它又产生睾丸激素(男性荷尔蒙),那么一些多用途的胚胎结构发展为阴茎和阴囊。但是如果他的睾丸激素接受器有缺陷(不是由性基因发展的),那么他的多用途胚胎结构将发展为阴蒂、阴唇和阴道,此时他的睾丸埋葬于腹股沟或阴唇之中了。这种假两性人常常是漂亮女孩,在女性模特中有许多案例呈现。
在实验室的实验中,如果雄性小鼠在出生或这之前阉割,成为成鼠后它们就不能显示雄性典型的性行为,甚至给它们注射雄性荷尔蒙也不行。但如果在出生几天以后再给它们阉割,成鼠后,雄性荷尔蒙能够引发雄性典型的行为。
综其上述,可以得到三个结论:
*一,那些早期事件确实影响或决定将来的结果。即,过去和将来有作用。
*二,那些行为(通过人类实验或通过基因自身)本身并不提供明确的结果。仅有合乎时宜的行为才能导致一个明确的结果。
*三,下游的行为也能够改变由早期行为导致的进程。
我们太多地认为这类体型发展是自然事件,就习以为常了。实际上,一个刚刚受精的卵子,就已经知道它的成体形态了,当它变成成体是应该是什么样子。这并不是因为那个受精卵是个预言家,也不是因为它是严格的基因决定论者。它仅仅遵循了过去和将来的作用(基因层次和环境作用)。
今天与将来有关联作用,这确实是事实。中国的独生子国策将决定(或改变)今后几十年的中国社会的人口结构。国会的财政预算的争论将决定美国社会的前景。过去也和今天和明天作用。孔子的话依旧主导每个中国人的思维过程。
但是,现在物理学不能解释明天在何处?昨天在哪里?明天如何变成今天的?今天如何和明天作用?我们肯定知道,明天并总是要变成今天。上述例子(生命和社会的发展)清楚地显示了今天确在和将来作用。
不能够回答上述问题,并不仅是因为当前主流物理尚未成熟,而且还因为我们未能真正地理解生命是什么。我发表于1993年莫斯科的19届全球哲学大会的论文—终极TOE(Theory of Everything,万物论),我已提供了一个说明明天在何处的模型。
V:相互包容无处不在
—生命及其和物理、数学、神学间联系的形而上学基础
在有丝分裂(染色体的复制)中,成千上万的基因代码被复制。用今天的技术和定量控制标准,在每百万仅产生200个缺陷的进程被认为是极可靠的进程。但如果在任何染色体复制中每十亿中仅有一个错,每个物种在10代内将失去它至关重要的基因特征。在果蝇例子中,它的复眼要求六个光接受细胞(观察空间一点)连接到相同的一对神经元。一项跟踪650个神经元的研究表明没有一个和细胞进行错误连接。
一方面,尽管人类大脑的发展确实是由基因控制的,但在这个过程中也会有许多的变动产生。即使是具有完全一样的基因的同卵双生子也不会有相同的神经系统。这是后成的一种表达。因此,每一个人在其历史上都记下了他或她的大脑的每一个结构。
因此,不仅仅基因决定论是有弹性的,而且在绝对可靠性结构中也有无限种的可能性。绝对可靠性保证了每一个人类胚胎(即便是有许多缺陷的基因)都可以成功地成长为人类。而这种无限可能性则保证了单个个体都是真实的。
此外,任何生命的发展,都只有一个过程 — 身体的发展。但这个单一的过程却产生了两个绝对相反的趋势。
- 从有序到无序,单个卵分化成为众多细胞。
- 从无序到有序,那些分化的细胞组织成为器官,从而成为组织得更好的身体。
然而,这两个相反的趋势在同一个过程中得到完成。事实上,想从单一的开始,建造成为组织得更好的结构的唯一方法,便是首先变为无序状态(分化)。也就是说,混沌是有序的原因和驱动力。简言之,无序和有序是事物的两个方面。相同的,绝对可靠性和无限可能性也是事物的两个方面。
这种两个绝对相对的趋势、力量或概念之间的相互关系称为互相共有 (互含)。这个概念,中国人早在5000年之前就已经用阴阳作出了表述。在有冲突的时候,阴阳是对立的两个方面,但长久而言,他们不但互补而且相互根植于对方当中。当达到一定的界限时,它们可以互相转化,也就是,阴变为阳,阳变为阴。因此,阴(弱的一方)可以通过屈服,从而征服阳(强的一方) 。使之达到最大值,最终使阳转化为阴。这个概念不仅是个强有力的政策战略,也是理解终极真理的唯一方法。
现代物理的基础是海森堡测不准原理的哥本哈根解释(CI)所表达的互补原理,它包括以下三个部分:
- 整体必须包含两个相反的部分。
- 这两个部分互相排斥。
- 这两个部分互补。
很显然,哥本哈根解释并没有反映出不确定性原理的真实含义。当我们想更精确地测量出一个粒子的位置时(ΔX变小),就必定失去了其动量的精确信息(ΔP变大)。也就是,正如阴–阳转化一样,ΔP转化为ΔX。ΔP和ΔX互相包含(互相共有),而非互相排斥。海森堡不确定性原理实际上是物理学中相互共有相互包容的原理。
VI:体形和大脑结构的规律
在果蝇实验中,当两条腿变大,而其它腿保持不变时,不同的片段一定是由不同的基因所控制的。当两条腿变肥而不是拉长时,不同的腿的轴也是被独立控制。表面上看起来,基因是控制体形的最佳候选。但是,如果我们比较人类和黑猩猩,就可以发现其中有超过99%的氨基酸序列是相同的。许多差异并不会产生功能性的结果,所以很难相信这些差异导致了两种物种的形态差异。更何况,有些基因非常保守,在长时间的演化中只会改变一点点,而有些则是飞速进化。然而,对这两种进化,快和慢的两组进行比较后发现,它们的蛋白质序列的改变速率几乎相同。具有完全不同的头部形状的两种果蝇(D. Heteroneura 和D. Sylvestries)可以异种交配。 D. W. Thompson发现通过在不同的轴中,简单地作出有比例的改变,可以实现两种形体之间的转化。他写道:“有一些东西…。不可缺少的…在这变形中…每一点每一条线…在整个变形过程中维持相对的顺序和位置…”
因此,尽管体形对自然选择的反应非常迅速,但进化过程还是要遵循几何规律。简单地说,几何才是体形的控制者,而基因只不过是个蓝领工作者。
人类的大脑中,有100亿个神经细胞(神经元)和10万亿多的连接(神经腱)。在大脑的发展过程中,神经元经历了剧烈的迁移活动,在这个迁移过程中,每个神经元发射出大约1000个树枝状结晶(短连接)和许多神经轴突(长连接) 。这样可以与其他的神经元发生联系,从而形成神经网络。神经原是怎么知道,何去何从呢?在大脑的三维空间中,又是怎么实现发射过程的?他们怎么从数十亿的细胞当中,找到欲与之联系的细胞呢?
单个的神经原,不可能拥有以上问题的所有知识和答案。大脑的发展只是一个神经原的剧烈迁移过程。那些没有建立正确联系的神经原都会死亡。在大脑的胚胎形成时,大约有10% 到80%(依区域而定)的神经原要死亡。
这个现象给出了一个非常重要的结论。早在神经原剧烈迁移之前,人脑就已有一个网络结构。那些迁移的神经原,面临的局面是, 不能嵌到预先的结构中去,便是死亡。这个预设的大脑网络, 是由拓扑结构描述。体形规程也是如此。
VII: 对称性及其对称性破缺
大部分的昆虫或动物都具有左右对称,而不是什么上下对称或前后对称。也许,上下不对称还可能由于引力而产生。那前后对称呢?确实,许多植物因为不能四处走动,它们拥有四个方向的对称。而动物则可以方便地走来走去。从生物学经济角度出发,便失去了前后对称。但这个对称的破坏却有着比生物经济更为深远的超自然原因。
在蛾中,皮屑(神经细胞)的发展、表皮细胞的不对称分化,给出了一个退化的内在更小的子细胞。外层细胞又偏斜地分裂,产生了成穴的外部细胞以及成皮的内部细胞。这个细胞的分化(对称的破坏)来自于一个母细胞的分化,这不可能是由基因的行为引起的,而是与拓扑关系有关。
对称和对称的破缺是物理学和数学的中心点,它们实际上产生了整个宇宙(时间、空间和质量)。对称性破缺产生所有的力!细胞的分化产生生命力!一层一层往上将追寻宇宙的起源,一层一层往下就是最基本的物理底层的电磁力。宇宙就是这样的一个统一体。